
14 Nov 2008
Many swimming bacteria are propelled by flagellar motors that stochastically switch between the clockwise and counterclockwise rotation direction. While the switching dynamics are one of the most important characteristics of flagellar motors, the mechanisms that control switching are poorly understood. We present a statistical-mechanical model of the flagellar rotary motor, which consists of a number of stator proteins that drive the rotation of a ring of rotor proteins, which in turn drives the rotation of a flagellar filament. At the heart of our model is the assumption that the rotor protein complex can exist in two conformational states corresponding to the two respective rotation directions, and that switching between these states depends on interactions with the stator proteins. This naturally couples the switching dynamics to the rotation dynamics, making the switch sensitive to torque and speed. Another key element of our model is that after a switching event, it takes time for the load to build up, due to polymorphic transitions of the filament. Our model predicts that this slow relaxation dynamics of the filament, in combination with the load dependence of the switching frequency, leads to a characteristic switching time, in agreement with recent observations.

05 Oct 2009
We demonstrate experimentally that disordered scattering can be used to
improve, rather than deteriorate, the focusing resolution of a lens. By using
wavefront shaping to compensate for scattering, light was focused to a spot as
small as one tenth of the diffraction limit of the lens. We show both
experimentally and theoretically that it is the scattering medium, rather than
the lens, that determines the width of the focus. Despite the disordered
propagation of the light, the profile of the focus was always exactly equal to
the theoretical best focus that we derived.
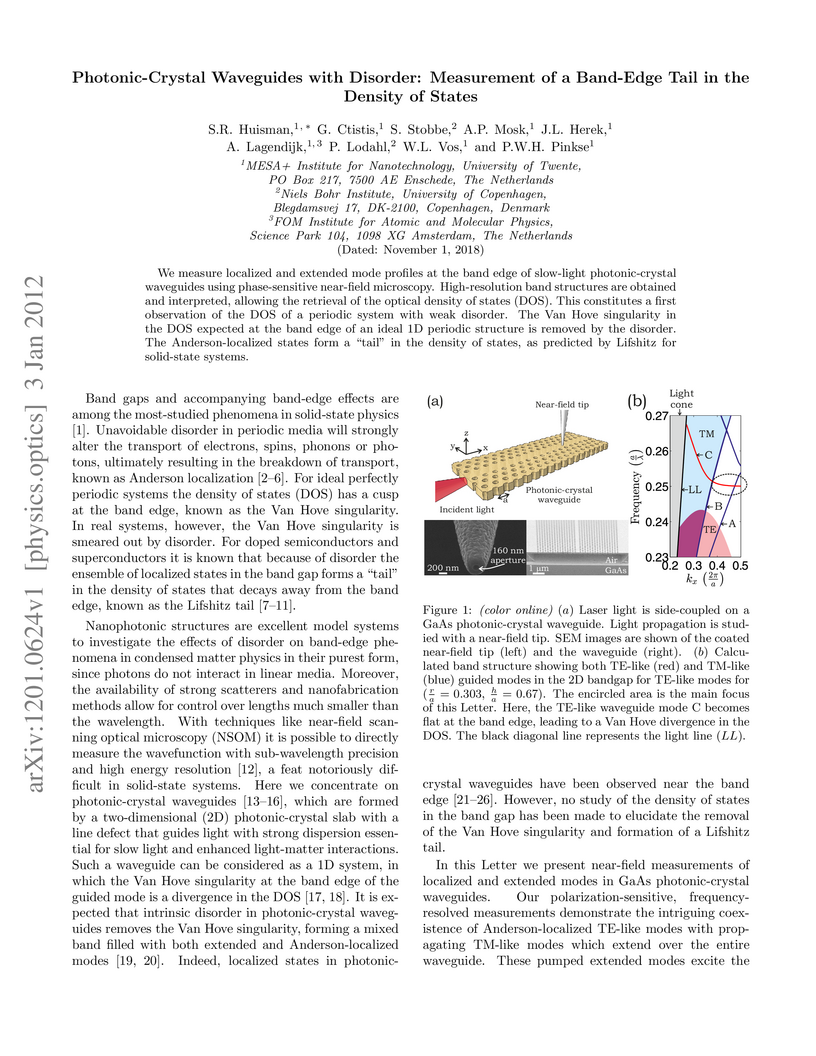
03 Jan 2012

We measure localized and extended mode profiles at the band edge of slow-light photonic-crystal waveguides using phase-sensitive near-field microscopy. High-resolution band structures are obtained and interpreted, allowing the retrieval of the optical density of states (DOS). This constitutes a first observation of the DOS of a periodic system with weak disorder. The Van Hove singularity in the DOS expected at the band edge of an ideal 1D periodic structure is removed by the disorder. The Anderson-localized states form a "tail" in the density of states, as predicted by Lifshitz for solid-state systems.

11 Dec 2009

We recently found that crystallization of monodisperse hard spheres from the bulk fluid faces a much higher free energy barrier in four than in three dimensions at equivalent supersaturation, due to the increased geometrical frustration between the simplex-based fluid order and the crystal [J.A. van Meel, D. Frenkel, and P. Charbonneau, Phys. Rev. E 79, 030201(R) (2009)]. Here, we analyze the microscopic contributions to the fluid-crystal interfacial free energy to understand how the barrier to crystallization changes with dimension. We find the barrier to grow with dimension and we identify the role of polydispersity in preventing crystal formation. The increased fluid stability allows us to study the jamming behavior in four, five, and six dimensions and compare our observations with two recent theories [C. Song, P. Wang, and H. A. Makse, Nature 453, 629 (2008); G. Parisi and F. Zamponi, Rev. Mod. Phys, in press (2009)].

12 Jul 2009
Diamonds melt at temperatures above 4000 K. There are no measurements of the steady-state rate of the reverse process: diamond nucleation from the melt, because experiments are difficult at these extreme temperatures and pressures. Using numerical simulations, we estimate the diamond nucleation rate and find that it increases by many orders of magnitude when the pressure is increased at constant supersaturation. The reason is that an increase in pressure changes the local coordination of carbon atoms from three-fold to four-fold. It turns out to be much easier to nucleate diamond in a four-fold coordinated liquid than in a liquid with three-fold coordination, because in the latter case the free-energy cost to create a diamond-liquid interface is higher. We speculate that this mechanism for nucleation control is relevant for crystallization in many network-forming liquids. On the basis of our calculations, we conclude that homogeneous diamond nucleation is likely in carbon-rich stars and unlikely in gaseous planets.

17 Sep 2002
We discuss 3-body processes in ultracold two-component Fermi gases with short-range intercomponent interaction characterized by a large and positive scattering length . It is found that in most cases the probability of 3-body recombination is a universal function of the mass ratio and , and is independent of short-range physics. We also calculate the scattering length corresponding to the atom-dimer interaction.

10 Mar 2005
We report a novel Monte Carlo scheme that greatly enhances the power of
parallel-tempering simulations. In this method, we boost the accumulation of
statistical averages by including information about all potential parallel
tempering trial moves, rather than just those trial moves that are accepted. As
a test, we compute the free-energy landscape for conformational changes in
simple model proteins. With the new technique, the sampled region of the
configurational space in which the free-energy landscape could be reliably
estimated, increases by a factor 20.

20 Dec 2012
NV centers in diamond are generally recognized as highly promising as indefinitely stable highly efficient single-photon sources. We report an experimental quantification of the brightness, radiative decay rate, nonradiative decay rate and quantum efficiency of single NV centers in diamond nanocrystals. Our experiments show that the commonly observed large spread in fluorescence decay rates of NV centers in nanodiamond is inconsistent with the common explanation of large nanophotonic mode-density variations in the ultra-small high-index crystals at near-unity quantum efficiency. We report that NV centers in 25 nm nanocrystals are essentially insensitive to local density of states (LDOS) variations that we induce at a dielectric interface by using liquids to vary the refractive index, and propose that quantum efficiencies in such nanocrystals are widely distributed between 0% and 20%. For single NV centers in larger 100 nm nanocrystals, we show that decay rate changes can be reversibly induced by nanomechanically approaching a mirror to change the LDOS. Using this scanning mirror method, for the first time we report calibrated quantum efficiencies of NV centers, and show that different but nominally identical nanocrystals have widely distributed quantum efficiencies between 10% and 90%. Our measurements imply that nanocrystals that are to be assembled into hybrid photonic structures for cavity QED should first be individually screened to assess fluorescence properties in detail.

04 Mar 2007


In a recent series of ground-breaking experiments, Nakajima et al. [Science
308, 414-415 (2005)] showed that the three cyanobacterial clock proteins KaiA,
KaiB, and KaiC are sufficient in vitro to generate circadian phosphorylation of
KaiC. Here, we present a mathematical model of the Kai system. At its heart is
the assumption that KaiC can exist in two conformational states, one favoring
phosphorylation and the other dephosphorylation. Each individual KaiC hexamer
then has a propensity to be phosphorylated in a cyclic manner. To generate
macroscopic oscillations, however, the phosphorylation cycles of the different
hexamers must be synchronized. We propose a novel synchronisation mechanism
based on differential affinity: KaiA stimulates KaiC phosphorylation, but the
limited supply of KaiA dimers binds preferentially to those KaiC hexamers that
are falling behind in the oscillation. KaiB sequesters KaiA and stabilizes the
dephosphorylating KaiC state. We show that our model can reproduce a wide range
of published data, including the observed insensitivity of the oscillation
period to variations in temperature, and that it makes nontrivial predictions
about the effects of varying the concentrations of the Kai proteins.

24 Sep 2009
Rare events are ubiquitous in many different fields, yet they are notoriously
difficult to simulate because few, if any, events are observed in a conventiona
l simulation run. Over the past several decades, specialised simulation methods
have been developed to overcome this problem. We review one recently-developed
class of such methods, known as Forward Flux Sampling. Forward Flux Sampling
uses a series of interfaces between the initial and final states to calculate
rate constants and generate transition paths, for rare events in equilibrium or
nonequilibrium systems with stochastic dynamics. This review draws together a
number of recent advances, summarizes several applications of the method and
highlights challenges that remain to be overcome.

03 Jul 2009

Multisite covalent modification of proteins is omnipresent in eukaryotic
cells. A well-known example is the mitogen-activated protein kinase (MAPK)
cascade, where in each layer of the cascade a protein is phosphorylated at two
sites. It has long been known that the response of a MAPK pathway strongly
depends on whether the enzymes that modify the protein act processively or
distributively: distributive mechanism, in which the enzyme molecules have to
release the substrate molecules in between the modification of the two sites,
can generate an ultrasensitive response and lead to hysteresis and bistability.
We study by Green's Function Reaction Dynamics, a stochastic scheme that makes
it possible to simulate biochemical networks at the particle level and in time
and space, a dual phosphorylation cycle in which the enzymes act according to a
distributive mechanism. We find that the response of this network can differ
dramatically from that predicted by a mean-field analysis based on the chemical
rate equations. In particular, rapid rebindings of the enzyme molecules to the
substrate molecules after modification of the first site can markedly speed up
the response, and lead to loss of ultrasensitivity and bistability. In essence,
rapid enzyme-substrate rebindings can turn a distributive mechanism into a
processive mechanism. We argue that slow ADP release by the enzymes can protect
the system against these rapid rebindings, thus enabling ultrasensitivity and
bistability.
There are no more papers matching your filters at the moment.
